TECIDO NERVOSO
O tecido nervoso é responsável por controlar as atividades dos órgãos e sistemas. É dividido anatomicamente em dois:
- Sistema nervoso central (SNC): formado pelo encéfalo e pela medula espinal. Ambas as estruturas revestidas por meninges, camadas de tecido conjuntivo circundantes. O córtex cerebral corresponde à camada mais externa do encéfalo, local onde ocorrem sinapses e os corpos celulares estão condensados, em cortes frescos, apresenta-se como uma substância escura. O oposto é encontrado no interior do encéfalo, onde há grande quantidade de fibras axônicas formando tratos, devido ao acúmulo de mielina, recebe o nome de substância branca. No entanto, no meio das fibras existem diversos corpos celulares, parecidos com pequenas ilhotas de substância cinzenta, são os núcleos. Já na medula espinal, o arranjo das substâncias é invertido, pois a substância branca ocupa a periferia da medula (no corte transversal) e o centro é composto pela substância cinzenta que forma uma espécie de ‘H’ ao redor do canal central. Os neurônios da coluna anterior da medula são motores, ao passo que os corpos de neurônios sensitivos estão agrupados nos gânglios dorsais e geralmente são neurônios pseudounipolares.
- Sistema nervoso periférico (SNP): formado pelos nervos cranianos, espinais e periféricos. Além dos gânglios, grupo de corpos celulares fora do SNC, e terminações nervosas que podem ser sensitivas ou motoras
A relação entre o SNC, os nervos sensitivos e motores cria as vias neurais, essas intermediam os arcos reflexos do organismo, reação raramente processada pelo encéfalo que interage por sinapses diretamente com os neurônios motores. Funcionalmente, a divisão também ocorre em dois grupos:
- Sistema nervoso somático (SNS): fornece inervação motora e sensitiva, controla as funções voluntárias e conscientes do organismo. Os arcos reflexos não fazem parte dessa divisão.
Sistema nervoso autônomo (SNA): fornece inervação motora involuntária e sensitiva das vísceras. A contração do músculo liso, controlado por este sistema, pode alterar o diâmetro dos órgãos ocos, das vísceras e vasos; nas glândulas, o SNA vai controlar a composição e secreção do epitélio glandular; e no sistema cardiovascular, cuja interferência do sistema autônomo pode alterara a frequência de despolarização do marca-passo cardíaco. Nesse grupo ainda há subcategorias:
- Sistema simpático
- Sistema parassimpático
- Divisão entérica
Os neurônios pré-sinápticos da divisão simpática estão localizados na porção torácica e lombar superior da medula espinal, ao passo que da divisão parassimpática estão localizados no tronco encefálico e na porção sacral. Geralmente, os neurônios pós-sinápticos parassimpáticos e sinápticos inervam as mesmas estruturas com ações antagonistas entre si.
Em várias partes do encéfalo, os neurônios atuam como células secretoras, esse tipo de tecido é denominado neuroendócrino, e deriva da cooperação entre o sistema nervoso e o sistema endócrino.
O neurônio é a célula funcional do sistema nervoso, que tem como função receber estímulos e transmitir para outra célula através de seus prolongamentos citoplasmáticos. A junção entre dois prolongamentos neuronais recebe o nome de sinapse. Auxiliando esse processo, existem as células de sustentação, chamadas de células da glia, mas não participam da transmissão elétrica. No SNC quatro grupos celulares fazem essa função: oligodendrócitos, astrócitos, micróglia e células ependimárias. Ao passo que no SNP temos células de Schwann e células-satélites. As funções das células da glia são: proteção física, remoção dos neurotransmissores das fendas sinápticas, isolamento dos corpos e prolongamentos dos neurônios, regulação do meio líquido do SNC e da interação entre o sistema circulatório e o tecido nervoso. Os numerosos vasos que irrigam tanto o SNC quanto o SNP são revestidos pela barreira hematoencefálica, que possui como função impedir o vazamento de substâncias dessas estruturas para o tecido nervoso, isto é, fazem o papel de filtração, evitando flutuações na concentração de eletrólitos e metabólitos que circulam pelos vasos do sistema nervoso. Essa barreira é formada pelos podócitos dos astrócitos nas zônulas de oclusão dos endotélios.
O neurônio, como unidade estrutural do tecido nervoso pode ser classificado de acordo com sua função:
- Neurônio motor: transmitem impulsos elétricos do SNC ou de gânglios para as células efetoras. Os neurônios motores podem ser somáticos ou viscerais, respectivamente voluntários e involuntários.
- Neurônio sensitivo: transmitem impulsos dos receptores para o SNC. Também possuem classificação quanto a funcionalidade, sendo fibras aferentes somáticas responsáveis pela inervação superficial do corpo, músculos e tendões; e as fibras sensitivas viscerais que inervam os órgãos internos, mucosas e vasos sanguíneos.
- Interneurônio: formam uma rede de comunicação entre neurônios motores e sensitivos. Mais de 99% dos neurônios do tecido nervoso estão dentro dessa classificação.
Morfologia do neurônio: ele é composto por pericário/soma/corpo celular, que abrange o núcleo e as organelas responsáveis pelo funcionamento celular. O citoplasma ao redor do núcleo, citoplasma perinuclear, apresenta uma grande quantidade de RE rugoso e ribossomos livres, que na MO aparecem como agregados denominados corpúsculos de Nissl. Entre o pericário e o axônio há uma estrutura formada pelo centro organizador de microtúbulos chamado de cone axônico, desprovido de outras organelas e permite diferenciar um dendrito de um axônio. Axônio é o maior prolongamento citoplasmático, o único que transmite os impulsos do corpo para os terminais, locais de sinapse, seja em outros neurônios, ou em células efetoras. Cada neurônio possui apenas um axônio e essas estruturas podem ser enormes. Abaixo do cone axônico, já no axônio propriamente dito, existe o segmento axônico inicial que filtra as proteínas e lipídios do citoplasma axonal. Dendritos são prolongamentos menores, mas de diâmetro maior, que emergem do pericário, recebem impulsos e transmitem para o corpo celular. Diferentemente dos axônios, não são mielinizados e formam ramificações enormes, as chamadas árvores dendríticas, que aumentam a superfície receptora da célula por meio de espinhos dendríticos compostos por filamentos de actina. A formação desses espinhos está intimamente associada com a neuroplasticidade e aprendizado. E a classificação neuronal pode ser baseada também na sua morfologia, isto é, na quantidade de prolongamentos que emergem do corpo celular:
- Multipolares: possuem um axônio e dois ou mais dendritos. Recebem os impulsos desses dendritos ou pela soma, e transmitem o sinal elétrico do axônio, uma estrutura condutora, até o terminal sináptico. A maioria dos neurônios com essa morfologia são neurônios motores ou interneurônios.
- Bipolares: apresentam um axônio e somente um dendrito. São muito mais raros que os multipolares, servindo como receptores dos sentidos especiais.
- Pseudounipolares: possuem apenas um ramo emergindo do corpo celular, que se bifurca em dois. Um ramo dendrítico periférico, faz o papel de receber os impulsos, e o ramo axônico central, transmissor da informação. Esse tipo celular deriva dos neurônios bipolares, que fusionam seus dois prolongamentos em um só. E a maioria desses exerce função sensitiva próxima ao SNC.
Apesar da concepção geral de uma baixa taxa mitótica dos neurônios, recentemente foram encontradas células-tronco neurais capazes de regenerar o tecido em regiões como o giro denteado do hipocampo e no bulbo olfatório, mas que também possuem capacidade migratória.
O transporte de substâncias produzidas no corpo celular ocorre por meio dos microtúbulos citoplasmáticos, processo denominado transporte neuronal. Esse tem como finalidade levar substâncias da soma para os terminais ou vice-versa. Duas vias principais são responsáveis pelo fenômeno:
- Transporte anterógrado: do corpo celular para os terminais axônicos. As cinesinas, proteínas motoras direcionadas para a extremidade positiva (voltadas para o terminal), movem vesículas através dos microtúbulos utilizando energia da hidrólise de ATP.
- Transporte retrógrado: dos terminais para a soma. As dineínas fazem esse papel, deslocam-se para a extremidade negativa.
Além disso, o sistema de transporte pode ser diferenciado de acordo com a velocidade de transmissão de informações:
- Sistema de transporte anterógrado lento: geralmente ocorre transporte de proteínas estruturais como a actina e outras proteínas metabólicas. 0,2 a 4mm/dia.
- Sistema de transporte rápido: de sentido duplo, é dividido em anterógrado rápido e retrógrado rápido. O primeiro transfere produtos do pericário, proteínas de baixo peso molecular, ao passo que o segundo leva os mesmos tipos de materiais que sofreram endocitose.
Sinapses: são contatos entre neurônios ou entre um neurônio e uma célula efetora. Na junção neuronal, ela pode ser classificada como:
- Axodendrítica: contato entre dendrito e axônio.
- Axossomática: contato entre um axônio e o pericário.
- Axoaxônica: contato entre dois axônios.
Geralmente, o axônio do neurônio pré-sináptico cruza com vários espinhos dendríticos, formando os botões de passagem, até sua terminação, onde ele se dilata e forma o botão terminal. Assim como diversas outras características desse tecido, as sinapses também são classificadas, em dois grupos de acordo com sua natureza:
- Sinapse química: ocorre com a liberação de uma substância química no terminal sináptico, um neurotransmissor. E é formada por alguns elementos clássicos:
- Botão pré-sináptico: dilatação da terminação nervosa, local onde há liberação de vesículas contendo neurotransmissores que entrarão em contato com a membrana do neurônio pós-sináptico.
- Fenda sináptica: espaço que o neurotransmissor deve atravessar para sair de uma célula e chegar até a outra.
- Membrana pós-sináptica: local de fixação dos neurotransmissores, é caracterizada por uma camada elétron-densa de inúmeros receptores.
- Sinapse elétrica: não necessitam de um neurotransmissor porque a união sináptica entre dois neurônios se dá por junções comunicantes que permitem a passagem de corrente da célula pré-sináptica para a pós-sináptica.
Ademais as sinapses podem ser classificadas de acordo com sua reação no neurônio pós-sináptico:
- Sinapses excitatórias: estimulam a abertura de canais catiônicos, elevadores do potencial de membrana até um limiar de ação que provoca a despolarização da membrana e a geração de um impulso nervoso.
- Sinapses inibitórias: estimulam os canais aniônicos, o que aumenta a diferença entre o potencial de repouso e o limiar de ação. Isso provoca a hiperpolarização e torna a membrana menos excitável, inibindo a geração do potencial de ação na célula.
Desse modo, o impulso nervoso deriva do somatório de efeitos de centenas sinapses, de modo que até a intensidade do impulso seja controlada pelo resultado desse fenômeno.
A despolarização do terminal sináptico provoca a abertura dos canais de cálcio dependentes de voltagem, responsáveis por estimular a exocitose das vesículas sinápticas ou pela liberação dos ligantes por porocitose. Os neurotransmissores podem ter efeitos diversos a depender de sua natureza ou do receptor em que atuará. São divididos em duas categorias:
- Ionotrópicos: canais iônicos, são alterados pela interação com o ligante, e provoca o influxo ou efluxo iônico na célula pós-sináptica.
- Metabotrópicos: receptores do tipo proteína G, traduzem sinais externos em sinais internos por esta via.
Os neurotransmissores mais comuns são:
- Acetilcolina: neurotransmissor atuante entre neurônios, ou entre neurônios e células efetoras do SNA. As células secretoras de ACh são os neurônios colinérgicos, assim como os receptores para esse ligante recebem o nome de receptores colinérgicos.
- Catecolaminas: neurotransmissores derivados do aminoácido tirosina. São eles: epinefrina, norepinefrina e dopamina. Neurônios secretores são catecolaminérgicos e atuam na parte motora, humoral e na atenção.
- Serotonina: deriva da triptofano e atua como neurotransmissor no SNC e na divisão entérica do SNA. Os neurônios secretores desse ligante são serotoninérgicos, e esse neurotransmissor já se mostrou crucial para o desenvolvimento assimétrico esquerdo-direito dos embriões.
- Aminoácidos: atuam principalmente no SNC. São exemplos: glicina, glutamato, aspartato e gama-aminobutírico.
- Óxido nítrico: sintetizado dentro da sinapse e utilizado imediatamente, sem a necessidade de síntese e armazenamento. Atua na via da proteína G.
A maioria dos neurotransmissores são recuperados ou degradados na junção sináptica. Sendo boa parte captada pela célula pré-sináptica (80%) e a quantidade remanescente é captada pela membrana pós-sináptica.
Células de sustentação do sistema nervoso: neuroglia:
Neuroglia periférica: composta principalmente por células de Schwann e células-satélites que variam de acordo com o tecido em questão. As células de Schwann são responsáveis pela produção da bainha de mielina, camada de lipídios que reveste o axônio, no sistema nervoso periférico. Durante a formação da bainha de mielina, mielinização, a membrana das células de Schwann apresenta três domínios distintos de acordo com o grau de revestimento, a membrana plasmática abaxônica, adaxônica e o mesaxônio, união entre essas duas partes polares da membrana que enrola-se no axônio por um movimento espiralado. A quantidade de lamelas formadas determina a espessura da bainha, entretanto, essa variável está sob controle do neurônio e não da célula de Schwann, e ocorre por meio do fator de crescimento neurorregulina. Segmento internodal é o trecho do axônio revestido pela bainha, marcando esses segmentos está o nó de Ranvier, uma ‘área nua’ cujo impulso elétrico é regenerado devido uma maior densidade de canais de Na+ .
Células-satélites: protegem as células neuronais, promovendo um microambiente controlado e uma via para trocas metabólicas.
Neuroglia central: dividida em quatro grupos celulares:
- Astrócitos: fornecem suporte físico e metabólico aos neurônios do SNC por meio do tamponamento especial do potássio, isto é, controlam a concentração de potássio extracelular no microambiente neuronal. Se ligam aos neurônios e modulam suas atividades, através de conexões que formam espécies de pés terminais entre os vasos sanguíneos e a célula neuronal. Ajudam também a manter a barreira hematoencefálica, controlar o trânsito de metabólitos do sangue para os neurônios e revestir os nódulos de Ranvier, evitando vazamento de neurotransmissores. São classificados em dois grupos de acordo com os prolongamentos citoplasmáticos:
- Astrócitos protoplasmático: encontrados na superfície encefálica, na substância cinza, possuem muitos prolongamentos curtos e ramificados.
- Astrócitos fibrosos: encontrados na substância branca, essas células apresentam menos prolongamentos.
- Oligodendrócitos: formação e manutenção da bainha de mielina somente no SNC. Se encontram alinhados entre fileiras de axônios, revestindo os espaços internodais. Diferentemente das células de Schwann, um oligodendrócito emite diversos prolongamentos, e cada prolongamento se enrola em um axônio, de modo que ele possa mielinizar mais de um neurônio simultaneamente. Portanto a razão entre essas células de isolamento elétrico e neurônios é menor no SNC do que no SNP.
- Células ependimárias: células que revestem o canal central da medula e os ventrículos cerebrais. Formam uma espécie de tecido epitelioide nas cavidades preenchidas por líquido. A superfície apical apresenta cílios e microvilosidades relacionadas com a absorção do líquido cerebroespinal. No terceiro ventrículo foram encontrados tipos especiais de células ependimárias, os tanicitos, que são carentes de cílios e relacionados com o controle dos metabólitos presentes no fluido cerebroespinal. Nos ventrículos, o revestimento ependimário juntamente com os capilares adjacentes formam o líquido dessas cavidades, essas estruturas são denominadas plexo corióideo.
- Micróglia: apresenta propriedades fagocíticas e atuam na limpeza do tecido nervoso. As células dessa classe fazem parte do sistema fagocitário mononuclear e entram no SNC através da corrente sanguínea. São as menores células da neuroglia e atuam no combate contra bactérias, células defeituosas e restos celulares.
Condução dos impulsos nervosos: ocorre, como dito outrora, pela somação de efeitos excitatórios e inibitórios presentes na membrana do neurônio, a condução elétrica ocorrerá se houver despolarização no cone axônico para que o sinal alcance os terminais pré-sinápticos. Após a chegada do impulso no segmento axônico inicial, ele é regenerado conforme passa pelos nódulos de Ranvier, numa espécie de condução saltatória com a reversão da voltagem nesses locais.
Associação do tecido conjuntivo com o tecido nervoso: os nervos periféricos são organizados por camadas de tecido conjuntivo:
- Endoneuro: formado por TC frouxo que reveste cada uma das fibras nervosas. As fibrilas colágenas seguem paralelas e revestem a fibra nervosa, de modo a revestir todo o comprimento do axônio. Além dos fibroblastos, as únicas células do tecido conjuntivo encontradas no endoneuro são os mastócitos e os macrófagos que atuam em casos de vigilância e reparo do tecido em casos de lesões.
- Perineuro: formado por TC especializado que reveste cada fascículo, e um fascículo é formado por fibras axônicas. Esse revestimento contribui para a formação de uma barreira metabolicamente ativa, a barreira hematoneural para manter o equilíbrio do meio iônico nas fibras nervosas. As células formadoras dessa barreira são diferenciadas, apresentam junções oclusivas que garantem a integridade da camada celular, disposição em camadas e a presença de lâmina basal confere-lhes características comuns de tecido epitelióide. Diferentemente do endoneuro, não há células do sistema imune.
- Epineuro: formado por TC denso não modelado, e é a camada mais externa, circundando os fascículos formados pelo perineuro. Além do tecido conjuntivo, essa camada apresenta um percentual lipídico considerável em nervos mais calibrosos, além da vascularização que penetra a estrutura e permite que o perineuro faça a filtração metabólica bidirecional, de uma camada vascularizada (epineuro) até a camada mais interna não vascularizada (endoneuro).
Além das camadas conjuntivas que revestem os nervos periféricos, existem outras 3 camadas próprias do revestimento neural para o sistema nervoso central:
- Dura-máter: folheto espesso de tecido conjuntivo denso.
- Aracnóide-máter: camada mais sensível adjacente a dura-máter, apresenta projeções semelhantes a teias que atingem a pia-máter revestem o espaço subaracnóideo.
- Pia-máter: túnica delicada que reveste o SNC.
Receptores aferentes do sistema nervoso sensitivo: receptores possuem a capacidade de gerar um impulso em resposta a determinado estímulo, e são classificados em:
- Exteroceptores: reagem aos estímulos externos do corpo.
- Enteroceptores: reagem aos estímulos internos, de distensão.
- Proprioceptores: também reagem aos estímulos internos, fornecendo informações acerca da posição, movimento e tônus muscular.
Regeneração do tecido nervoso: pode ocorrer no SNP, mas raramente no SNC, uma vez que a barreira hematoencefálica impede a entrada de macrófagos no local da lesão e os oligodendrócitos e células da micróglia são incapazes de fagocitar o axônio degenerado. Essa etapa de fagocitose é crucial, pois a regeneração neural depende dela para ocorrer. A porção lesionada é degradada devido sua incapacidade de realizar o transporte axônico, esse fenômeno recebe o nome de degeneração anterógrado. Ocorre, primeiramente, a desintegração granular do axônio, fragmentação do citoesqueleto do axônio. Quando o axônio perde contato com as células de Schwann, elas secretam fatores de crescimento glial, que vai estimular a proliferação do axônio. Aliado a isso, os macrófagos ativados começam a fagocitar os restos de mielina para que o novo axônio possa crescer no local. No SNC não há uma remoção de mielina eficiente, isso estimula o processo de formação de uma cicatriz glial, ou gliose, cujos astrócitos se proliferam e ocupam o local de lesão axonal. Alguns pontos são relevantes nesse reparo, primeiro, se a lesão ocorrer muito próxima do pericário, a degeneração retrógrada na porção proximal pode comprometer a célula como um todo, e uma vez que uma fibra do SNP é seccionada, o músculo inervado por ela sofre atrofia. A informação recebida da degeneração retrógrada no corpo celular leva o neurônio a produzir novos prolongamentos nervosos, neuritos, que formam um cone de crescimento. Esse cone cresce e se desloca para a porção distal, isto é, para as terminações, e esse crescimento é guiado por células de Schwann desdiferenciadas que formam tubos para a passagem dos neuritos formadores do cone de crescimento, esse s tubos são denominados tubos endoneurais e se organizam longitudinalmente em bandas, chamadas bandas de Büngner. Assim que o contato do nervo motor for reestabelecido com o músculo, a contração muscular pode ocorrer novamente.
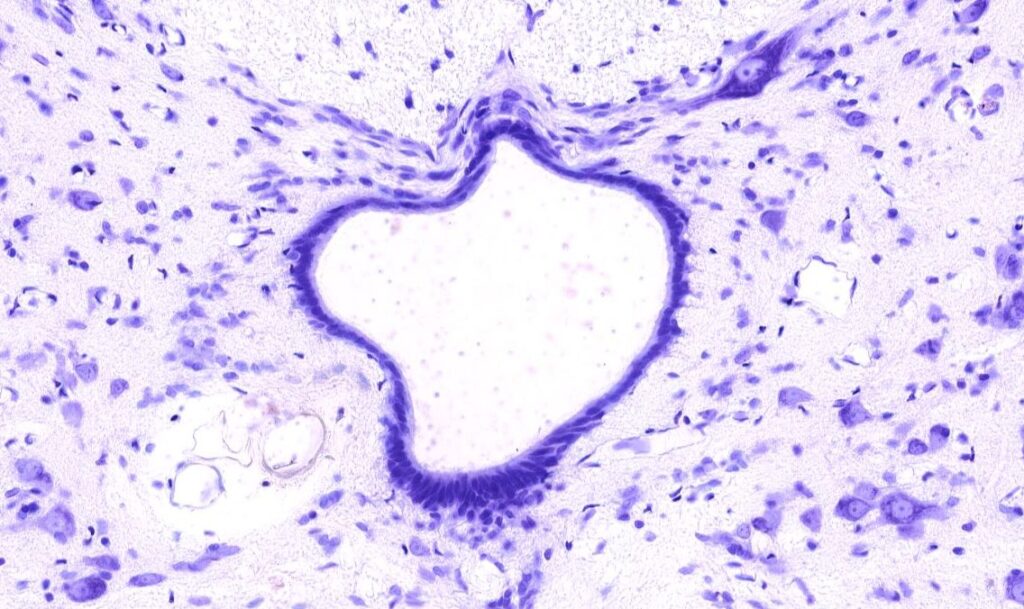
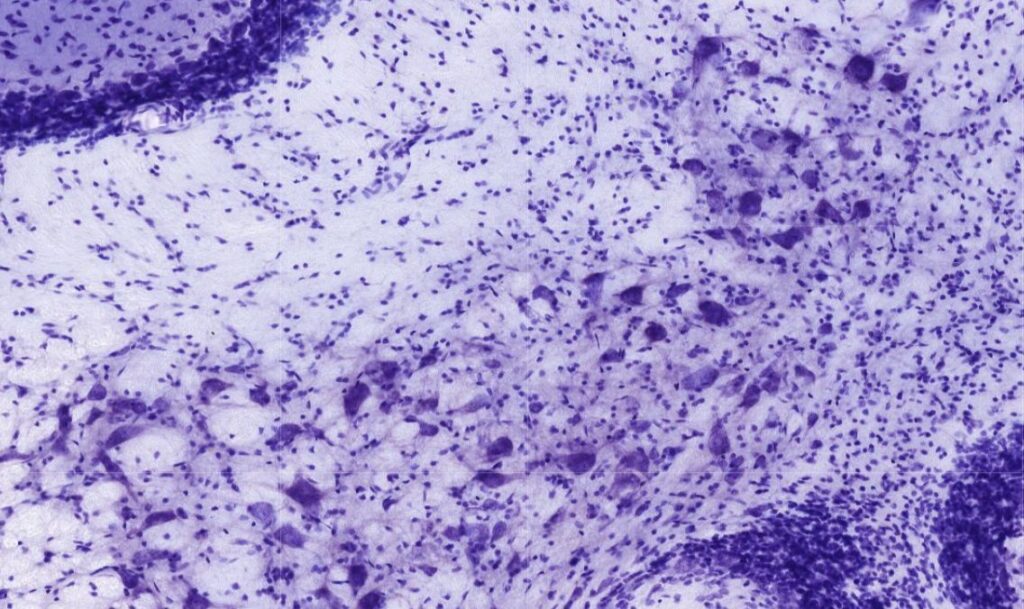
Todas as lâminas presentes no site são de autoria do site “https://histologyguide.com/“.
Universidade Estadual do Tocantins – UNITINS
2024