SISTEMA REPRODUTOR FEMININO
O sistema genital feminino é constituído por órgãos internos e externos. Os órgãos internos ficam na pelve e são os ovários, as tubas uterinas, o útero e a vagina. A genitália externa, também conhecida como pudendo feminino (vulva), inclui o monte do púbis, os lábios maiores e menores do pudendo, clitóris, o vestíbulo e abertura da vagina, o hímen e o óstio externo da uretra.
Ovário
O ovário tem como principais funções a produção de gametas e a de hormônios esteroides. Na gametogênese feminina, os gametas em desenvolvimento são chamados de oócitos, e os maduros, de óvulos. O ovário é responsável por produzir os estrogênios e os progestógenos. Os estrogênios promovem o crescimento e a maturação dos órgãos sexuais, além de contribuir para o desenvolvimento das mamas, enquanto os progestógenos prepararam os órgãos sexuais internos para a gravidez.
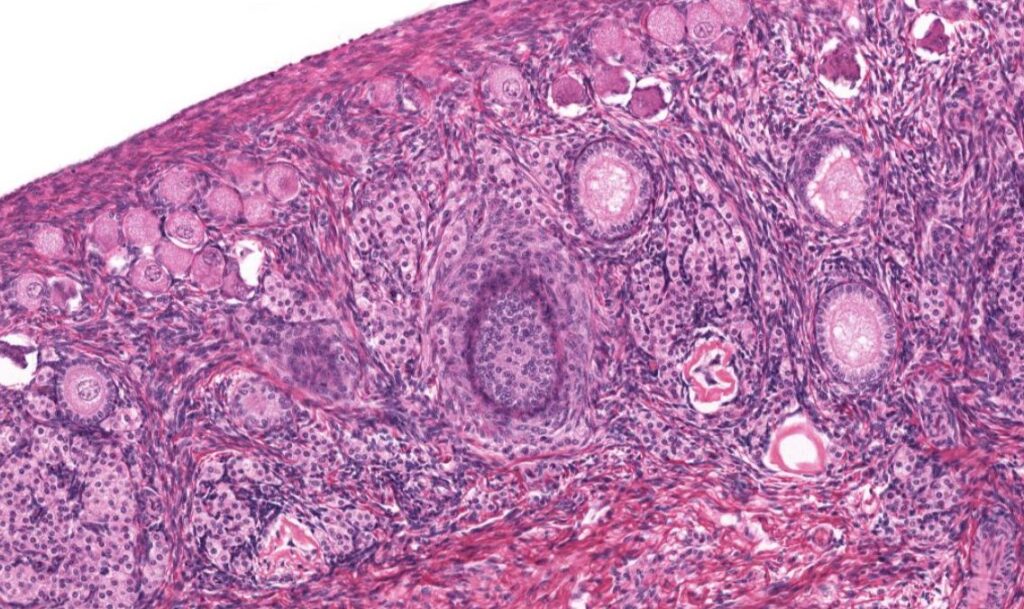
Os ovários consistem em um par de estruturas em formato de amêndoas, são fixados na superfície posterior do ligamento largo pelo mesovário. O polo superior do ovário é fixado pelo ligamento suspensor do ovário, e o polo inferior é ligado ao útero pelo ligamento útero-ovárico. O ovário é composto por uma medula e um córtex. A medula fica na parte central e contém tecido conjuntivo frouxo, vasos sanguíneos, vasos linfáticos e nervos. O córtex, localizado na parte periférica, abriga os folículos ovarianos entre tecido conjuntivo celularizado e fibras musculares lisas dispersas no estroma, envolvendo os folículos. Na superfície do ovário, há uma massa de células cuboides que formam o epitélio germinativo, essa camada é contínua com o mesotélio. Entre o epitélio germinativo e o córtex subjacente tem-se a túnica albugínea.
No estroma do córtex, encontram-se folículos de vários tamanhos. Durante a puberdade, pequenos grupos de folículos sofrem crescimento e maturação cíclicos, mas normalmente apenas um oócito é liberado do ovário a cada ovulação.
O desenvolvimento dos folículos é dividido em três estágios: folículos primordiais, folículos em crescimento e folículos maduros ou de Graaf. O folículo primordial é o estágio inicial, surgindo no terceiro mês do desenvolvimento fetal, e é encontrado no estroma do córtex. Nesse estágio, o oócito é circundado por uma camada de células foliculares pavimentosas, com a superfície externa dessas células delimitada por uma lâmina basal. O oócito no folículo primordial tem cerca de 30 μm de diâmetro, com um grande núcleo e vários nucléolos dispersos. O citoplasma do oócito, chamado de ooplasma, contém o corpúsculo de Balbiani, estrutura com acúmulo de membranas e vesículas de Golgi, centríolos, mitocôndrias e lisossomos, além de lamelas anulares, estruturas semelhantes a pilhas porosas.
Após o estágio de folículo primordial, há o folículo primário, o primeiro estado do folículo em crescimento. Nessa etapa, o oócito aumenta de tamanho e ocorre a proliferação das células foliculares circundantes, que se tornam cuboides. Além dessas células, forma-se a zona pelúcida, um revestimento extracelular formado pela secreção de proteínas pelo oócito. A zona pelúcida, localizada entre o oócito e as células foliculares adjacentes, contém glicoproteínas responsáveis por induzir a reação acrossômica.
No folículo primário também ocorre a formação da membrana granulosa pela rápida reprodução mitótica das células foliculares, que circunda o oócito. Nessa fase, as camadas da teca são formadas pelas células do tecido conjuntivo. As células do estroma que circundam o folículo formam a teca folicular, localizada externamente à lâmina basal e diferenciada em teca interna e teca externa. A teca interna é altamente vascularizada e composta por células secretoras cuboides, que produzem esteroides e possuem receptores de hormônio luteinizante (LH), de modo que, produzem e secretam androgênios em resposta à estimulação do LH. A teca externa é composta por células musculares lisas e feixes de fibras colágenas. A lâmina basal que separa a teca interna da camada granulosa, restringindo a entrada de leucócitos e substâncias de alto peso molecular no folículo.
Durante o estágio do folículo primário, ocorre a maturação do oócito, com a formação dos grânulos corticais, vesículas secretoras de proteases no momento em que o espermatozoide ativa o óvulo. O oócito também forma microvilosidades no espaço perivitelino, além do desenvolvimento de prolongamentos das células da granulosa para o oócito, que formam junções comunicantes com a membrana plasmática do oócito. As células da granulosa produzem monofosfato cíclico de guanosina (cGMP), que entra no oócito pelas junções comunicantes e inibe a hidrólise de monofosfato cíclico de adenosina (cAMP) por fosfodiesterase (PDR3A), bloqueando a progressão da meiose I. Além disso, o peptídeo natriurético do tipo C (CNP), secretado pelas células da granulosa, atua como inibidor da continuação da meiose, podendo interromper a meiose por um período de 12 a 50 anos.
Posteriormente, o folículo primário aumenta de tamanho devido à proliferação das células da granulosa, até a formação de 6 a 12 camadas de células. Cavidades com líquido surgem entre essas células, que coalescem até formar uma cavidade única, o antro, atingido o estágio de folículo secundário ou antral. Nessa fase, é formada a coroa radiada pelas células da granulosa.
Por fim, tem-se o folículo maduro ou de Graaf, com o oócito secundário maduro. Esse folículo possui cerca de 10 mm, nesse estágio, o estágio granuloso torna-se mais fino. O oócito e as células do cúmulo são afastados, e as camadas da teca tornam-se mais proeminentes.
A produção de estrógenos no ovário depende da interação entre as células da teca interna e da granulosa. Uma vez que as células da teca interna são estimuladas pelo LH, convertendo colesterol em pregnenolona e, após, em androgênios. Entretanto, as células da teca interna não possuem a enzima P450 aromatase para produzir estrógenos, mas as células da granulosa têm essa enzima, de modo que os androgênios secretados pela teca interna são convertidos em estrógenos em resposta ao FSH. Posteriormente, tem-se o surto de LH que dessensibiliza os receptores de LH, fazendo as células da granulosa pararem de produzir estrogênios, o que reinicia a primeira divisão meiótica do oócito primário, formando o oócito secundário e o primeiro corpúsculo polar. As células da granulosa e da teca sofrem luteinização e passam a produzir de progesterona.
A ovulação é o processo que resulta na liberação do oócito secundário, ocorrendo no décimo quarto dia de um ciclo de 28 dias. Antes da ovulação, o fluxo sanguíneo é interrompido na área próxima ao folículo maduro, conhecida como mácula pelúcida, que eleva-se e rompe-se. Por conseguinte, o oócito circundado pela coroa radiada e pelas células do cúmulo do oóforo é liberado, então, a massa do cúmulo contendo o oócito é direcionada pelas fímbrias para o óstio abdominal da tuba uterina. O oócito permanece viável por 24 horas, porém, caso não ocorra a fertilização, o oócito sofre degeneração. Após o oócito ser liberado, a segunda divisão meiótica progride, mas é interrompida na metáfase, completando-se apenas se houver fertilização, formando um óvulo maduro e o segundo corpúsculo polar.
Depois da ovulação, ocorre a luteinização, processo no qual a parede folicular, composta de células da granulosa e tecais remanescentes, é transformada no corpo lúteo. As células aumentam de tamanho e se enchem de gotículas lipídicas, adquirindo uma aparência amarelada. As células derivadas da granulosa, que são grandes e localizadas centralmente, constituem a maioria do corpo lúteo e produzem estrogênio, progesterona e inibina. Enquanto as células provenientes da teca são menores e mais intensamente coradas, ficam na parte periférica e secretam androgênios e progesterona. Durante a formação do corpo lúteo, os vasos da teca interna crescem na granulosa e formam uma rica rede vascular. O corpo lúteo secreta progesterona e estrogênio para estimular o crescimento e revestimento do útero, o endométrio, para a implantação do zigoto. Caso não ocorra a fertilização, o corpo lúteo permanece ativo por 10 dias e, na ausência de gonadotropina coriônica humana, a secreção de progesterona e estrogênio é reduzida, levando à degeneração do corpo lúteo, formando uma cicatriz branca, o corpo albicans. Caso ocorra a fertilização, o corpo lúteo constituirá o corpo lúteo da gravidez, que depende das luteotropinas (estrogênios, IGF-I, IGF-II, HCG, LH e a insulina). O corpo lúteo produz progesterona, que bloqueia o desenvolvimento cíclico dos folículos ovarianos. O corpo lúteo da gravidez começa a ter sua função reduzida depois de 8 semanas de gestação, quando a placenta assume a sua função de produzir progesterona e estrógenos.
A maioria dos folículos sofre atresia para degenerar e desaparecer. Essa atresia é mediada pela apoptose das células da granulosa.
O suprimento sanguíneo dos ovários é dado principalmente pelas artérias ováricas, que se anastomosam-se com os ramos ováricos das artérias uterinas. As veias acompanham as artérias e formam o plexo pampiniforme, a partir do qual é formada a veia ovárica.
Os ovários são inervados pelo plexo ovárico autonômico.
Tubas Uterinas
As tubas uterinas são um par de tubos que ligam o útero aos ovários. Elas têm a função de transportar o óvulo do ovário até o útero e proporcionar um ambiente favorável para a fertilização e desenvolvimento inicial do zigoto. As tubas uterinas têm cerca de 10 a 12 cm de comprimento e são compostas por quatro partes: o infundíbulo, em formato de funil e adjacente ao ovário, onde se encontram as fímbrias, prolongamentos franjados que vão do óstio do infundíbulo até o ovário; a ampola, o maior segmento das tubas uterinas, onde ocorre a fertilização; o istmo, a parte medial da tuba uterina adjacente ao útero; e a parte uterina das tubas uterinas, que tem cerca de 1 cm de comprimento e fica na parede do útero.
A parede uterina é dividida em três partes: uma camada serosa externa, composta de mesotélio e de tecido conjuntivo; uma camada muscular intermediária; e uma camada mucosa interna com finas pregas que se projetam para o lúmen da tuba uterina. O revestimento mucoso contém epitélio simples colunar com células ciliadas e não ciliadas. As células ciliadas são mais numerosas na ampola e no infundíbulo, direcionando a onda dos cílios para o útero, enquanto as células não ciliadas secretam líquido nutritivo ao óvulo.
Útero
O útero fica localizado na pelve entre a bexiga e o reto, e é o local que recebe a mórula em desenvolvimento das tubas uterinas, onde ocorre o desenvolvimento embrionário e fetal. O útero tem 7,5 cm de comprimento, 5 cm de largura e 2,5 cm de espessura e é dividido em duas regiões: o corpo, que é a parte superior do útero, sendo que a sua parte superior e arredondada é chamada fundo do útero; e o colo, que é a parte inferior do útero. O colo do útero e o corpo do útero são separados pelo istmo. Além disso, o útero tem o óstio interno, que comunica-se com a cavidade do útero e o óstio externo, que comunica-se com a vagina.
A parede uterina é composta por três camadas: o endométrio, que é a mucosa do útero; o miométrio, que é a camada muscular espessa do útero; e o perimétrio, que é a camada serosa externa do útero. Essa camada mais externa é composta por mesotélio, uma camada fina de tecido conjuntivo frouxo e, abaixo do mesotélio, tem-se tecido elástico. O perimétrio não recobre totalmente a face anterior do útero, sendo que essa região é recoberta por tecido conjuntivo ou adventícia.
O miométrio tem três camadas de músculo liso: o estrato vascular, camada muscular média, sendo a mais espessa; a camada externa e a camada interna. Essas camadas atuam como um sincício funcional durante a contração desse órgão. Em comparação com o corpo do útero, o colo do útero apresenta menos tecido muscular e mais tecido conjuntivo.
Em relação à estrutura do endométrio, tem-se o estrato funcional, que é a parte espessa do endométrio que sofre descamação durante a menstruação e também tem-se a camada basal, que permanece durante a menstruação. O endométrio também é revestido por epitélio simples colunar com células secretoras e ciliadas. Este epitélio de superfície se invagina na lâmina própria subjacente, o estroma endometrial, formando as glândulas uterinas. Essas glândulas são tubulares simples, contendo menos células ciliadas e se ramificam na camada profunda do endométrio. Esse estroma endotelial é muito celularizado e possui uma quantidade abundante de matriz extracelular. Em relação à vasculatura do endométrio, essa também prolifera e degenera no decorrer do ciclo menstrual.
Outrossim, endométrio e o miométrio sofrem alterações durante o ciclo, que são divididas em fase proliferativa, secretora e menstrual. A fase proliferativa é regulada pelos estrogênios e nesse período, as células estromais, endoteliais e epiteliais do estrato basal se proliferam. Após, tem-se a fase secretora que é regulada pela progesterona, nesse processo, o endométrio torna-se edematoso, aumentando a sua espessura, as glândulas aumentam de tamanho e o líquido mucoide produzido pelo epitélio glandular é rico em nutrientes responsáveis pela nutrição do embrião em caso de implantação, também tem-se a hipertrofia das células epiteliais e aumento da vascularização. Também tem-se a diferenciação das células do estroma em células deciduais, ricas em glicogênio que proporcionam a nutrição do embrião. Por fim, tem-se a fase menstrual, resultante do declínio de progesterona e de estrogênio. Primeiramente, na fase menstrual, ocorrem contrações periódicas das paredes das artérias espiraladas, o que vai ocasionar isquemia do estrato funcional, as glândulas do endométrio diminuem a secreção e a espessura do endométrio é reduzida, consequentemente, ocorre a ruptura o epitélio superficial e dos vasos sanguíneos, de modo que a descamação do endométrio continue até que se tenha somente o estrato basal.
Caso ocorra a fecundação, a fase menstrual é substituída pela fase gravídica e, na segunda semana, tem-se a fixação do cório e as células da placenta começam a secretar hCG e outras luteotropinas, que mantêm o corpo lúteo ativo e a produção de progesterona e estrogênios, esses hormônios evitam a involução do endométrio e estimula o seu desenvolvimento nas primeiras semanas da gravidez. Antes de ser implantado, o blastocisto fica livre no lúmen uterino e possui uma massa externa denominada trofoblasto e uma massa interna denominada embrioblasto. No processo de implantação, o trofoblasto começa a invadir o endométrio e a parte invasora diferencia-se em sinciciotrofoblasto e citotrofoblasto. Após 11 dias de desenvolvimento, o blastocisto fica totalmente inserido no endométrio. Depois da implantação, o endométrio sofre decidualização, quando os fibroblastos do estroma endometrial diferenciam-se em células deciduais em consequência à progesterona. A decídua possui três regiões diferentes: a decídua basal subjacente ao local de implantação, a decídua capsular entre o local de implantação e o lúmen do útero e a decídua parietal que inclui o endométrio remanescente.
A mucosa cervical, diferentemente do restante do útero, possui grandes glândulas ramificadas. Além disso, o endométrio do colo do útero sofre pouca alteração da sua espessura no decorrer do ciclo menstrual. Ademais, as glândulas cervicais sofrem alterações funcionais importantes, uma vez que na metade do ciclo a quantidade de muco aumenta 10 vezes, sendo que o muco é menos viscoso, proporcionando um ambiente favorável para a passagem dos espermatozoides.
Placenta
A placenta é formada por uma parte fetal (cório) e uma parte materna (decídua basal) e está relacionada com a troca de substâncias entre as circulações materna e fetal. Essa circulação é formada pelo desenvolvimento das vilosidades coriônicas. O crescimento da placenta é realizado pelo crescimento intersticial da concha trofoblástica. Também tem-se células mesenquimatosas, células reticulares, fibroblastos, miofibroblastos, células musculares lisas e células apresentadoras de antígenos placentárias fetais, estas últimas participam de reações imunes inatas placentárias.
No início do desenvolvimento, os espaços intervilosos são locais para a troca de nutrientes, produtos metabólicos, e resíduos , porém, à medida que o crescimento prossegue, as vilosidades da decídua capsular começam a degenerar, produzindo uma superfície lisa, o cório liso. Também tem-se a formação do cório viloso, altamente ramificado com vilosidades adjacentes à decídua basal. De modo que a camada da placenta a partir da qual as vilosidades se projetam é denominada placa coriônica.
Do quarto ao quinto mês de gestação, a porção fetal da placenta é dividida pelos septos placentários em cotilédones, onde o sangue materno pode circular. A decídua basal forma o componente materno da placenta e a placa basal é a parte externa da placenta que está em contato com a parede uterina.
Ademais, o sangue materno e o sangue fetal são separados pela barreira placentária, mantida pelas camadas de tecido fetal, essas camadas são muito finas para facilitar a troca de substâncias e é formada por sinciciotrofoblasto, camada citotrofoblástica interna delgada, lâmina basal do trofoblasto, tecido conjuntivo da vilosidade, lâmina basal do endotélio e endotélio do capilar placentário fetal da vilosidade terciária.
Além do mais, a placenta consiste em um local de trocas gasosas, de metabólitos e de anticorpos entre as circulações materna e fetal, essas substâncias são trocadas através das finas camadas fetais que separam as duas correntes sanguíneas. Essa troca ocorre à medida que o sangue flui pelas vilosidades. O sangue materno irriga a placenta por meio de artérias espiraladas endometriais que penetram a placa basal.
Além disso, a placenta também é considerada um órgão endócrino, pois produz progesterona, essencial para a manutenção da gravidez; estrogênio; gonadotropina coriônica humana (hCG), necessária para a implantação e manutenção da gravidez; somatomamotropina coriônica humana (hCS), promotora do crescimento, reguladora da glicose e estimuladora da proliferação dos ductos mamários; o IGF-I e o IGF-II estimulam a proliferação e diferenciação do citotrofoblasto; fator de crescimento endotelial (EGF); relaxina, amolece o colo do útero e os ligamentos pélvicos; e a leptina, regula a reserva de nutrientes maternos e o transporte desses nutrientes para o feto através da barreira placentária.
Vagina
A vagina consiste em um tubo fibromuscular que liga os órgãos genitais internos ao ambiente externo, estendendo-se do colo do útero até o vestíbulo. A abertura da vagina pode ser circundada pelo hímen, pregas da mucosa derivadas da membrana endodérmica. A parede da vagina é composta por uma camada mucosa interna, uma camada muscular intermediária e uma camada adventícia externa. A camada mucosa interna é revestida por epitélio estratificado pavimentoso, contém numerosas pregas transversais e possui papilas de tecido conjuntivo projetando-se para a camada epitelial. A camada muscular possui uma camada longitudinal externa e uma camada circular interna. Por fim, a camada adventícia contém uma camada interna de tecido conjuntivo denso com numerosas fibras elásticas e uma camada externa de tecido conjuntivo frouxo com nervos, vasos sanguíneos e linfáticos.
Além disso, o epitélio da vagina é desprovido de glândulas e sofre alterações cíclicas durante o ciclo menstrual. De modo que, durante a fase folicular, as células acumulam glicogênio, porém, na fase menstrual, a camada superficial do epitélio a vagina pode sofrer descamação.
Genitália Externa
A genitália externa consiste em um conjunto de estruturas denominado pudendo feminino com epitélio estratificado pavimentoso. A genitália externa é composta pelo monte do púbis, proeminência arredondada acima da sínfise púbica, composta por tecido adiposo subcutâneo; pelos lábios maiores do pudendo, grandes pregas longitudinais da pele, com uma camada fina de músculo liso, grande quantidade de tecido adiposo subcutâneo e glândulas sebáceas e sudoríparas; lábios menores do pudendo, um par de pregas de pele, que margeiam o vestíbulo, possui vasos sanguíneos e fibras elásticas delgadas, além de glândulas sebáceas no estroma; o clitóris é uma estrutura homóloga ao pênis, composta por dois pequenos corpos cavernosos, glande, dois ramos do clitóris e o prepúcio com inúmeras terminações nervosas; tem-se também o vestíbulo da vagina que é revestido por epitélio estratificado, possui numerosas glândulas mucosas.
Glândulas Mamárias
As glândulas mamárias desenvolvem-se na puberdade sob a influência dos hormônios estrogênio e progesterona, sofre um aumento de tamanho devido ao crescimento do tecido adiposo interlobular e os ductos se estendem e se ramificam dentro do estroma do tecido conjuntivo em expansão.
Mesmo após a puberdade, as glândulas mamárias ficam inativas até a gravidez, quando os estrogênios, progesterona, a prolactina e os gonadocorticoides promovem a maturação dessas glândulas. Além disso, a mudança hormonal da menopausa involui a glândula mamária, sendo substituída por tecido adiposo e conjuntivo.
As glândulas mamárias são sudoríparas apócrinas tubuloalveolares modificadas, sendo que a glândula mamária adulta inativa é composta de 15 a 20 lobos intercalados por faixas fibrosas de tecido conjuntivo, esses lobos irradiam-se a partir da papila mamária e são subdivididos em unidades lobulares do ducto terminal (ULDT), algumas faixas fibrosas, denominadas ligamentos suspensores, conectam–se com a derme. Também tem-se uma quantidade abundante de tecido adiposo no tecido conjuntivo denso dos espaços interlobulares. Cada glândula termina em um ducto galactóforo, que se abre através de um orifício contraído na papila mamária. Abaixo da aréola, cada ducto dilata-se no seio galactóforo e, perto de suas aberturas, os ductos são revestidos por epitélio estratificado pavimentoso queratinizado. Ademais, a epiderme da papila mamária e da aréola é coberta por epitélio estratificado pavimentoso queratinizado. Abaixo da aréola e da papila mamária tem-se feixes de fibras musculares lisas dispostas no tecido conjuntivo denso. A aréola contém glândulas sebáceas, sudoríparas e mamárias.
A ramificação dos ductos galactóforos leva à unidade lobular do ducto terminal (ULDT), que representa um agrupamento de pequenos alvéolos, que forma um lóbulo com ductos terminais, ducto coletor intralobular e estroma intralobular, sendo que as células epiteliais glandulares e as células mioepiteliais constituem as mais importantes relacionadas aos dúctulos e lóbulos mamários.
Ademais, a morfologia da porção secretora da glândula mamária varia com o ciclo menstrual. De modo que na glândula inativa o componente glandular é esparso e consiste, principalmente, em ductos. Na fase folicular o estroma intralobular é menos denso e os dúctulos terminais são formados por células epiteliais cuboides com pouco lúmen . Na fase lútea, as células epiteliais ficam mais altas e o lúmen pode ser visto pelo acúmulo de secreções. Nos últimos dias do ciclo, tem-se involução e apoptose.
Durante a gravidez, as glândulas mamárias proliferam, de modo que, no primeiro trimestre tem-se o alongamento e ramificação dos dúctulos terminais e a proliferação das células epiteliais de revestimento e das células mioepiteliais. No segundo trimestre, tem-se a diferenciação dos alvéolos, o estroma do tecido conjuntivo intralobular é infiltrado por plasmócitos, linfócitos e eosinófilos, a quantidade de tecido glandular e a massa da mama aumentam de tamanho devido ao crescimento dos alvéolos. No terceiro trimestre, as células glandulares epiteliais tornam-se cuboides e possuem retículo endoplasmático rugoso extenso, vesículas secretoras e gotículas de lipídios, tem-se o declínio da proliferação das células estromais interlobulares e o aumento da mama pela hipertrofia das células secretoras e do acúmulo de produto secretor nos alvéolos.
Na produção de leite, tanto a secreção merócrina quanto a apócrina estão envolvidas, de modo que na secreção merócrina, o componente proteico do leite é sintetizado no RER, armazenado em vesículas e liberados por fusão da membrana da vesícula com a membrana plasmática. Na secreção apócrina, o componente de gordura ou lipídio do leite surge como gotículas de lipídios livres no citoplasma, o lipídio coalesce para formar gotículas de lipídios maiores, que vão para a região apical da célula e projetam-se para o lúmen do ácino.
Lorem ipsum dolor sit amet, consectetur adipiscing elit. Ut elit tellus, luctus nec ullamcorper mattis, pulvinar dapibus leo.

folículos primordiais: localizados no córtex externo do ovário.
Todas as lâminas presentes no site são de autoria do site “https://histologyguide.com/“.
Universidade Estadual do Tocantins – UNITINS
2024